

 :""гЊбјбјжГЪЕбщКѓГЃГЃЭЈЙ§ВЁОњИаШОЪЕбщРДЦРЙРгуЬхЕФНЁПЕ(LimЕШ,2009).ЪЕМЪбјжГжаГЃМћЕФЪШЫЎЦјЕЅАћОњОГЃБЛгУдкгЊбјУтвпЪЕбщжа,ИаШОЪШЫЎЦјЕЅАћОњКѓЛсГіЯжГібЊадАмбЊжЂзДМАРЃбёадВЁБф.дкЪЕМЪбјжГжаЪШЫЎЦјЕЅАћОњИаШОвВЪЧЕЫЎгубјжГИпЫРЭіТЪЕФдвђжЎвЛ,дьГЩСЫОоДѓЕФО",:::"жаЙњЫЎВњбјжГЭј",::"ЕЫЎВњЦЗ Внгу",
:""гЊбјбјжГЪЕбщКѓГЃГЃЭЈЙ§ВЁОњИаШОЪЕбщРДЦРЙРгуЬхЕФНЁПЕ(LimЕШ,2009).ЪЕМЪбјжГжаГЃМћЕФЪШЫЎЦјЕЅАћОњОГЃБЛгУдкгЊбјУтвпЪЕбщжа,ИаШОЪШЫЎЦјЕЅАћОњКѓЛсГіЯжГібЊадАмбЊжЂзДМАРЃбёадВЁБф.дкЪЕМЪбјжГжаЪШЫЎЦјЕЅАћОњИаШОвВЪЧЕЫЎгубјжГИпЫРЭіТЪЕФдвђжЎвЛ,дьГЩСЫОоДѓЕФО",:::"жаЙњЫЎВњбјжГЭј",::"ЕЫЎВњЦЗ Внгу",еЊвЊЃК
БОЪЕбщжМдкЦРЙРЫЧСЯжЌжЪЫЎЦНЖдВнгугзгуЩњГЄЁЂЗЧЬивьадУтвпМАИЮдрбѕЛЏзДЬЌЕФгАЯьЁЃдк27.5ЁР0.5ЁцЫЎЮТЯТгкЫЎзхЯфбЛЗЯЕЭГжагУ5жжВЛЭЌжЌжЪЫЎЦНЃЈ0ЁЂ25ЁЂ50ЁЂ75КЭ100 g/kgЫЧСЯЃЌЦфжагугЭ/гёУзгЭ=1:1ЃЉЕФДПЛЏЫЧСЯЮЙбјГѕЪМЬхжиЮЊ6.57ЁР0.01 gЕФВнгугзгу10жмЁЃНсЙћБэУїЃЌ50 g/kgЫЧСЯжЌЗОзщЕФдіжиТЪзюИпЃЌЮожЌЖдеезщзюЕЭЃЛЫЧСЯаЇТЪЁЂЕААзжЪаЇТЪКЭдіжиТЪЕФБфЛЏФЃЪНвЛжТЃЛгыЮожЌЖдеезщЯрБШЃЌЦфЫќИїзщЕФЗЧЬивьадУтвпжИБъЪ§жЕНЯКУЃЈШчЭЬЪЩЯИАћЛюадЁЂбЊНЌЙ§бѕЮяУИЛюадКЭШмОњУИЛюадЃЉЧвИЮдрбѕЛЏГЬЖШНЯЕЭЃЌЕЋЪЧЙ§СПЕФЫЧСЯжЌжЪЫЎЦНЛсЖдИЮдрдьГЩДњаЛИКЕЃЁЃдкбјжГЪЕбщНсЪјКѓЃЌЖдВнгуНјааСЫЪШЫЎЦјЕЅАћОњЃЈAeromonas hydrophilaЃЉИаШОЪЕбщЁЃНсЙћЯдЪОЃЌЮожЌЖдеезщВнгуДцЛюТЪзюЕЭЃЛ75 g/kgЫЧСЯзщВнгуДцЛюТЪзюИпЃЌЯджјИпгкЮожЌЖдеезщЃЈP <0.05ЃЉЁЃБОЪЕбщбаОПБэУїЪЪЕБЕФЫЧСЯжЌжЪКЌСПгажњгкЬсИпВнгуУтвпФмСІМАЯИОњИаШОЯТЕФДцЛюТЪЃЌЕЋЪЧЫЧСЯжЌжЪЙ§СПЛсдіМгВнгуИЮдрбѕЛЏГЬЖШЁЃдіжиТЪЕФЖўДЮЛиЙщЧњЯпЗжЮіБэУї6.6-35.5 gДѓаЁЕФВнгузюЪЪЫЧСЯжЌжЪЫЎЦНЮЊ65.3 g/kgЁЃ
1ЁЂЧАбд
гЩгкНќФъРДЫЧСЯвЕЕФХюВЊЗЂеЙМАбаОПШЫдБЕФВЛаИХЌСІЃЌФПЧАВнгуЩЬвЕЫЧСЯвбЛљБОПЩвдТњзуВнгуПьЫйЩњГЄЕФгЊбјашвЊЁЃШЛЖјВЛТлЪЧШ§ДѓгЊбјЫиЛЙЪЧЮЂСПгЊбјЫиЕФашЧѓСПбаОПЖМЪЧвдЩњГЄЧщПіЛђепЪЧЗёЕМжТШБЗІжЂЮЊБъзМЃЈLimЕШЃЌ2011ЃЉЃЌЖјВЛЪЧвдНЁПЕжИБъЮЊВЮПМЃЌАќРЈУтвпгІД№КЭМВВЁЕжПЙЁЃФПЧАЦЕЗББЌЗЂЕФМВВЁе§бЯжиЭўаВзХИпУмЖШИпВњСПбјжГФЃЪНЕФЗЂеЙЁЃУПФъгЩгкЪШЫЎЦјЕЅАћОњЃЈAeromonas hydrophilaЃЉЕФИаШОЃЌжаЙњВнгубјжГЫ№ЪЇДяЪ§АйЭђдЊЁЃ
ЭЈЙ§гЊбјВЙИјРДМѕЧсМВВЁЖдЫЎВњбјжГгАЯьЕФЙлФювбОдНРДдНБЛДѓМвНгЪмЃЈViswanathЃЌ2012ЃЉЁЃгЊбјЦНКтЃЌгШЦфЪЧдкзагуМАгзгуНзЖЮЃЌЖдЩњГЄЁЂПЙВЁМАДцЛюЖМгазХЩюдЖЕФгАЯьЃЈViswanathЃЌ2012ЃЉЁЃжЌжЪзїЮЊЫЧСЯжажївЊЕФФмСПРДдДЃЌгУгкТњзубјжГЦЗжжЕФФмСПМАБиашжЌЗОЫсашЧѓЁЃбаОПепдкЖўЪЎЪРМЭдчЦкЗЂЯжЩуЪГжЌжЪЖдЮЌГжКуЮТЖЏЮяЕФНЁПЕжСЙиживЊЃЈZibohЃЌ2000ЃЉЃЌДЫКѓдкБфЮТЖЏЮявВгаЯрЭЌЗЂЯжЃЈTakeuchiЕШЃЌ1977ЃЉЁЃвдЭљЕФбаОПжївЊМЏжадкВЛЭЌжЌЗОдДЛђепБиашжЌЗОЫсЖдУтвпгІД№ЛђПЙВЁФмСІЕФгАЯьЃЈLimЕШЃЌ2008ЃЉЃЌШЛЖјЫЧСЯжЌжЪЫЎЦНгыгуРрНЁПЕжЎМфЕФЙиЯЕЫљЪмЕФЙизЂЯрЖдНЯЩйЃЈLimЕШЃЌ2009ЃЉЁЃБОЪЕбщжМдкЦРЙРЫЧСЯжЌжЪЫЎЦНЖдВнгугзгуЩњГЄЁЂИЮПЙбѕЛЏадМАЗЧЬивьадУтвпгІД№ЕФгАЯьЁЃ
2ЁЂВФСЯгыЗНЗЈ
2.1 ЪЕбщЫЧСЯ
БОЪЕбщЙВЩшМЦСЫ5жжВЛЭЌжЌЗОКЌСПЕФЫЧСЯЃЌЪЕбщЫЧСЯХфЗНКЭГЩЗжЗжЮіШчБэ1ЫљЪОЁЃL0зщЮЊЮожЌЖдеезщЃЌL1ЃЌL2ЃЌL3ЃЌL4зщжажЌЗОЬэМгЫЎЦНЗжБ№ЮЊ25 g/kgЃЌ50 g/kgЃЌ75 g/kgЃЌ100 g/kgЁЃЪЕбщЫЧСЯдкГЃЮТЯТЗчИЩ24аЁЪБКѓжУ-20ЁцБљЯфБЃДцЁЃ
2.2 ЪЕбщгубјжГКЭЫЧбјЙмРэ
ВнгуШЁздБОЪЕбщЪвгцГЁЃЌбјжГЪЕбщПЊЪМЧАНЋгудкЪвФкбЛЗЫЎзхЯфжаднбј2жмвдЪЪгІЪЕбщЛЗОГКЭВйзїЃЌднбјЦкМфЭЖЮЙЫЧСЯL2ЁЃе§ЪНЪЕбщПЊЪМКѓЃЌУПЯфЃЈ200 LЃЉЫцЛњЗХгу30ЮВЃЌГѕЪМЬхжиЮЊ6.57ЁР0.01gЁЃУПзщЫЧСЯЩшШ§ИіЦНааЯфЃЌАДгуЬхжиЕФ4%~5%ШЗЖЈУПШеЭЖЫЧСПЃЌЗжБ№гк9ЁУ00КЭ16ЁУ00ИїЭЖЮЙвЛДЮВЂЙлВьгуЩњГЄЧщПіЃЌМЧТМЫРЭіЧщПіЁЃУП2жмГЦвЛДЮзмЬхжиЃЌВЂЯргІЕїећЭЖЮЙСПЁЃЫЧбјжмЦкЮЊ10жмЁЃЪЕбщЦкЫЎЮТЮЊ27.5ЁР0.5ЁцЃЌЫЎжаШмбѕСП7.5ЁР0.43 mg/LЃЌpHжЕЮЊ7.4ЁР0.2ЃЌАБЕЊЮЊ0.079ЁР0.005 mg/LЃЌУПШеЙтееЪБМфЮЊ12 hЁЃ
2.3 бљЦЗВЩМЏКЭЬхГЩЗжЗжЮі
ЪЕбщНсЪјКѓЃЌНЋгуНћЪГ24hКѓЃЌећЯфгуГЦжиЁЂЧхЕуЮВЪ§ЁЃУПЯфЫцЛњШЁгу3ЮВЙЉШЋгугЊбјГЩЗжЗжЮігУЃЌСэШЁ6ЮВЃЌТщзэКѓЗжБ№ГЦжиЃЌВтСПЬхжиЁЂЬхГЄЃЌЪЙгУОЙ§ИЮЫиФЦДІРэЙ§ЕФзЂЩфЦїДгЮЇаФЧЛШЁбЊЃЌСЂМД4ЁцРыаФЃЈ4000 r/minЃЌ10ЗжжгЃЉЃЌШЁЩЯВубЊНЌВЂжУгк-80ЁцБљЯфБЃДцЃЛдйАбгуНтЦЪШЁЕУФкдрЃЌЗжРыИЮдрКЭГІЯЕФЄжЌЗОЃЌЗжБ№ГЦжиМЧТМЃЌВЂМєШЁБГВПСНВрАзМЁЁЃ
ЫЧСЯМАбљЦЗЫЎЗжЁЂДжЕААзЁЂДжжЌЗОКЭЛвЗжЗжБ№ВЩгУ105ЁцГЃбЙИЩдяЗЈЁЂПЪЯЖЈЕЊЗЈЁЂЫїЪЯГщЬсЗЈКЭ550ЁцзЦЩеЗЈВтЖЈЁЃбЊНЌИЪгЭШ§ѕЅЁЂЕЈЙЬДММАМюадСзЫсУИЃЈALPЃЉЁЂЙШВнзЊАБУИЃЈASTЃЉКЭЙШБћзЊАБУИЃЈALTЃЉЛюаддкШ§ЬьФкВЩгУбЊвКздЖЏЩњЛЏЗжЮівЧКЭЯрЙиЪдМСКаВтЖЈЁЃ
КєЮќБЌЗЂЕФВтЖЈВЩгУNBTЗЈЃЌвдОоЪЩЯИАћВњЩњЕФбѕЛЏздгЩЛљВњЮяЃЈROSЃЉРДБэЪОЁЃбЊНЌШмОњУИЛюадЁЂбѕЛЏУИЃЈPODЃЉКЭИЮдрГЌбѕЛЏЮяЦчЛЏУИЃЈSODЃЉЁЂБћЖўШЉЃЈMDAЃЉМАзмПЙбѕЛЏФмСІЃЈT-AOCЃЉЛюадВЩгУЪдМСКаВтЖЈЁЃ
2.4 ЯИОњгІМЄЪЕбщ
ЪШЫЎЦјЕЅАћОњЃЈAhЃЌBSK-10ЃЉАДAlexanderЕШЃЈ2011ЃЉЕФЗНЗЈЛюЛЏСНДЮЁЃЪЕбщШЁбљНсЪјвЛжмКѓЃЌУПЯфШЁ10ЬѕгуИЙЧЛзЂЩф0.1 mLЕФОњаќИЁвКЃЈLC50ЃЌ6ЁС108 CFU/mLЃЉЁЃСэШЁ10ЬѕВнгуИЙЧЛзЂЩфЭЌЕШМССПЕФPBSЛКГхвКзіЮЊИКЖдеезщЁЃзЂЩфКѓе§ГЃЭЖЮЙЃЌдкНгЯТРДЕФЮхЬьМЧТМУПЬьИїзщгуЫРЭіЧщПіЃЌВЂШЗШЯЫРЭідвђЁЃЕкСљЬьМЦЫуДцЛюТЪЁЃ
2.5 ЭГМЦЗжЮі
ЪЕбщНсЙћгУЦНОљЪ§ЁРБъзМЮѓБэЪОЃЌОANOVAЗжЮіКѓЃЌгУDuncanЁЏsЖржиБШНЯЗЈЃЈDuncanЃЌ1955ЃЉЗжЮіИїДІРэМфЕФВювьЯджјадЃЈP <0.05ЃЉЁЃЫљВЩгУШэМўЮЊSPSS13.0ЁЃ
3ЁЂНсЙћ
3.1 ЩњГЄЁЂЫЧСЯРћгУЁЂЬхзщГЩМАбЊНЌУИЗжЮі
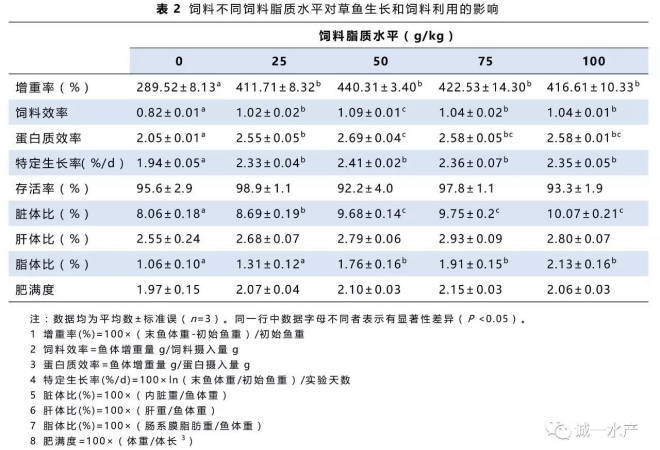
ЩуЪГВЛЭЌЫЧСЯВнгуЕФдіжиТЪЁЂЫЧСЯаЇТЪЁЂЕААзжЪаЇТЪМАДцЛюТЪШчБэ2ЫљЪОЁЃЭЖЮЙВЛЭЌжЌжЪЫЎЦНЕФЫЧСЯЖдДцЛюТЪУЛгаЯджјадгАЯьЃЈP >0.05ЃЉЁЃ50 g/kgЫЧСЯжЌжЪЫЎЦНзщЕФВнгудіжиТЪзюИпЃЌЦфДЮЪЧ25 g/kgЃЌ75 g/kgКЭ100 g/kgЫЧСЯжЌжЪЫЎЦНзщЃЌЮожЌЖдеезщЕФдіжиТЪЯджјЕЭгкЦфЫќИїзщЃЈP <0.05ЃЉЁЃЫЧСЯаЇТЪМАЕААзжЪаЇТЪЕФБфЛЏЧїЪЦКЭдіжиТЪЯрЭЌЁЃ
50 g/kgЁЂ75 g/kgКЭ100 g/kgЫЧСЯжЌжЪЫЎЦНзщВнгудрЬхБШКЭжЌЬхБШЯджјИпгкЦфЫќИїзщЃЌЖјЮожЌЖдеезщзюЕЭЁЃЫцзХЫЧСЯжЌжЪЫЎЦНЕФЬсИпЃЌИЮЬхБШгыЗЪТњЖШОљГЪЯжЩЯЩ§ЧїЪЦЃЌЕЋЮоЯджјадВювьЁЃ
ШчБэ3ЫљЪОЃЌЖдеезщКЭ25 g/kgЫЧСЯжЌжЪЫЎЦНзщШЋгуЫЎЗжКЭЛвЗжЖМИпгкЦфЫќИїзщЃЌ100 g/kgЫЧСЯжЌжЪЫЎЦНзщзюЕЭЁЃЯрЗДЃЌЫцзХЫЧСЯжЌжЪЫЎЦНЕФЩ§ИпЃЌШЋгужЌЗОКЌСПЯджјдіМгЃЌ100 g/kgЫЧСЯжЌжЪЫЎЦНзщЕФШЋгужЌЗОКЌСПЯджјИпгкЦфЫќИїзщЁЃИїзщМфМЁШтЫЎЗжгыжЌЗОЕФБфЛЏгыШЋгуЯрЫЦЁЃИїзщМфЕФШЋгуЕААзОљУЛгаЯджјВювьЁЃ
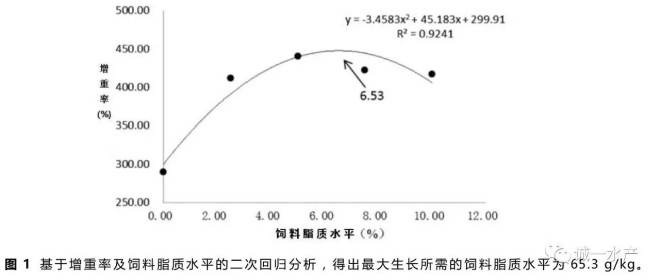
ИљОндіжиТЪМАЫЧСЯжЌжЪЫЎЦНЃЌЭЈЙ§ЖўДЮЛиЙщЗНГЬЪНЃЌМЦЫуЕУЕНВнгуЫЧСЯжЌжЪашЧѓСПЮЊ65.3 g/kgЃЈМћЭМ1ЃЉЁЃЖўЯюЪНЙЋЪНЮЊЃКy=-3.4583x2+45.183x+299.91ЃЌR2=0.9241ЁЃ

ЪЕбщИїзщВнгубЊНЌжаЕЈЙЬДМЁЂИЪгЭШ§ѕЅМАЯрЙиУИЛюЗжЮіМћБэ4ЁЃЕЭЫЧСЯжЌжЪЩуШызщЕФВнгубЊНЌжаМюадСзЫсУИЁЂЙШБћзЊАБУИКЭЙШВнзЊАБУИЛюадНЯИпЃЌЕЋЪЧИЪгЭШ§ѕЅМАЕЈЙЬДМКЌСПНЯЕЭЁЃВнгубЊНЌЕЈЙЬДМКЌСПЫцзХЫЧСЯжЌжЪЫЎЦНЩ§ИпгаЩЯЩ§ЧїЪЦЃЌЖјИЪгЭШ§ѕЅКЌСПдђЫцзХжЌЗОЩуШыЕФЬсИпЖјЯджјЩЯЩ§ЁЃМюадСзЫсУИЕФЛюадГЪЯжЯТНЕЕФЧїЪЦЁЃЙШБћзЊАБУИКЭЙШВнзЊАБУИЕФЛюадГЪЯжЯШНЕКѓЩ§ЕФЧїЪЦЃЌзюЕЭжЕГіЯждк50~75 g/kgЫЧСЯжЌжЪЫЎЦНзщЁЃ
3.2 ИЮдрЪмЫ№ЧщПіВтЖЈ
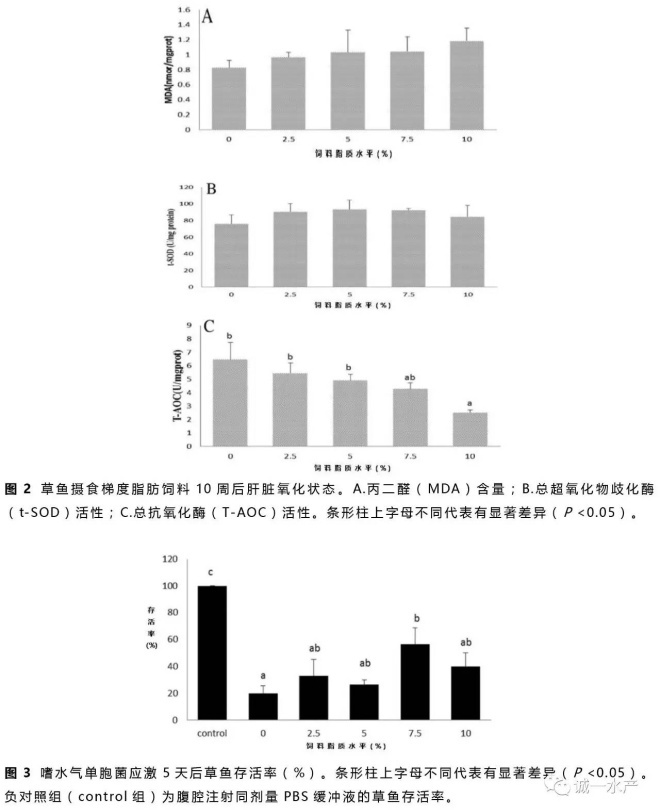
ИЮдрзмГЌбѕЦчЛЏУИЃЈt-SODЃЉЛюадЁЂБћЖўШЉЃЈMDAЃЉКЌСПМАзмПЙбѕЛЏУИЃЈT-AOCЃЉЛюадЗжЮіНсЙћМћЭМ2ЁЃИЮдрMDAКЌСПЫцзХЫЧСЯжЌжЪКЌСПдіМгЖјдіМгЃЈЭМ2AЃЉЃЌt-SODЛюадИїзщМфВювьВЛЯджјЃЈP >0.05ЃЌЭМ2BЃЉЃЌЕЋЪЧЩуЪГ50 g/kgЫЧСЯжЌжЪЕФВнгузщt-SODЛюадТдИпгкЦфЫќИїзщЁЃT-AOCЛюадЫцзХЫЧСЯжЌжЪКЌСПДг50 g/kgЩ§ЕН100 g/kgЖјЯджјНЕЕЭЃЈP <0.05ЃЉЃЌЕЋдкЕЭгк50 g/kgЫЧСЯжЌжЪКЌСПЕФИїзщМфВювьВЛЯджјЃЈP >0.05ЃЌЭМ2CЃЉЁЃ
3.3 ЪШЫЎЦјЕЅАћОњгІМЄКѓДцЛюТЪ
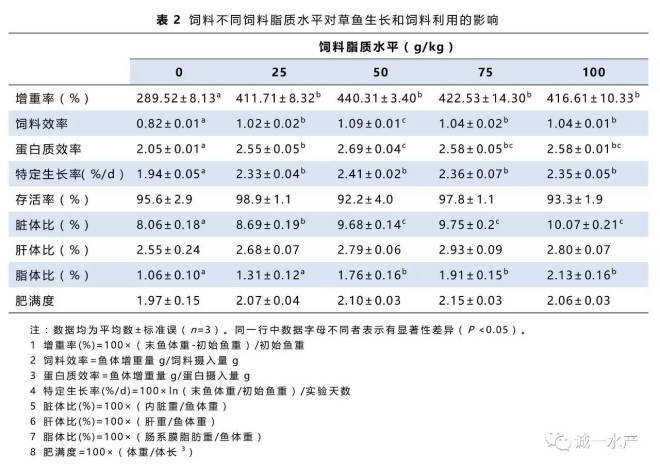
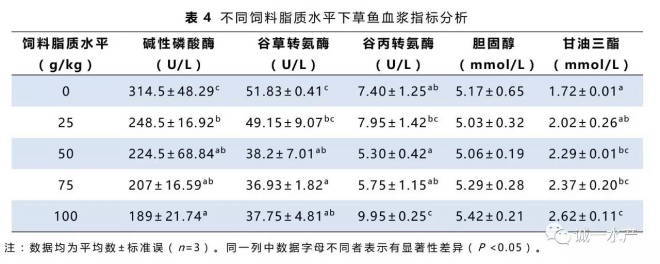
ВнгуИЙЧЛзЂЩфЪШЫЎЦјЕЅАћОњКѓЪзДЮЫРЭіГіЯждк12аЁЪБКѓЁЃ60аЁЪБКѓЫРЭіТЪЧїгкКуЖЈЁЃЫљгаЫРЭігуЖМГіЯжСЫРУЮВКЭРУШњМАГібЊадАмбЊжЂЯжЯѓЃЈRobertsЕШЃЌ1997ЃЉЁЃЪЕбщМЧТМСЫзЂЩфОњКѓ5ЬьФкЕФРлЛ§ЫРЭіТЪЃЌВЂдкЕкСљЬьМЦЫуДцЛюТЪЃЌНсЙћШчЭМ3ЫљЪОЁЃДцЛюТЪзюИпзщЮЊЩуЪГ75 g/kgЫЧСЯжЌжЪЕФВнгузщЁЃ0 g/kgжЌЗОзщЕФВнгуДцЛюТЪзюЕЭЃЈP <0.05ЃЉЁЃ

3.4 ВнгузЂЩфЪШЫЎЦјЕЅАћОњЧАКѓЕФЗЧЬивьадУтвпгІД№Люад
ЗЧУтвпгІД№ШчЭЬЪЩЛюадЁЂбЊНЌЙ§бѕЛЏЮяУИЃЈPODЃЉЛюадМАбЊНЌШмОњУИЛюадШчЭМ4ЫљЪОЁЃИаШОЧАЃЌЭЬЪЩЛюадЫцзХЫЧСЯжЌжЪЫЎЦНЕФЬсИпЖјГіЯжЩЯЩ§ЧїЪЦЃЌдк75 g/kgЫЧСЯжЌжЪзщГіЯжзюИпЗхжЕЁЃИаШОКѓЃЌЭЬЪЩЛюадЯджјНЕЕЭжСгІМЄЧАвЛАыЕФЫЎЦНЃЈЭМ4AЃЉЁЃ0 g/kgЫЧСЯжЌжЪзщЕФPODЛюадЯджјИпгкЦфЫќИїзщЃЈP <0.05ЃЉЃЛИаШОКѓЃЌИїзщPODЛюадЖМЯджјИпгкЦфгІМЄЧАЫЎЦНЃЈЭМ4BЃЉЁЃЫЧСЯжЌжЪЕФдіМгЯджјЬсИпСЫВнгубЊНЌШмОњУИЛюадЃЈP <0.05ЃЉЃЌИаШОКѓШмОњУИЛюаджЛЪЧТдЮЂгаЫљЩЯЩ§ЃЈЭМ4CЃЉЁЃ
4ЁЂЬжТл
4.1 ЫЧСЯжЌжЪЫЎЦНгыЩњГЄ
БОЪЕбщжаЃЌЩуЪГКЌжЌЫЧСЯЕФВнгудіжиТЪЯджјИпгкЮожЌЖдеезщЃЈP <0.05ЃЉЃЌгыжЎЧАвЛаЉбаОПБЈЕРЕФдіМгЫЧСЯжЌжЪКЌСПЛсДйНјгуРрЩњГЄЕФНсТлвЛжТЃЈWatanabeЕШЃЌ1982ЃЉЁЃЫЧСЯжЌжЪКЌСПгыВнгуЩњГЄЕФЙиЯЕЭЈЙ§ЖўЯюЪНЛиЙщЗжЮіШчЭМ1ЫљЪОЃЌзюДѓжЕГіЯждкЫЧСЯжЌжЪКЌСПЮЊ65.3 g/kgЕФЪБКђЁЃжЕЕУзЂвтЕФЪЧ25 g/kgЫЧСЯжЌжЪзщЕФВнгудіжиТЪгы50 g/kgЁЂ75 g/kgКЭ100 g/kgЫЧСЯжЌжЪзщЕФВнгудіжиТЪУЛгаЯджјВювьЃЈБэ2ЃЉЃЌЫЕУїдкЫЧСЯжЌжЪКЌСПДяЕН25 g/kgКѓвбОТњзуСЫВнгузюЕЭжЌЗОашЧѓСПЁЃБОЪЕбщжазюМбВнгуЫЧСЯжЌжЪКЌСПИпгкDuЕШЃЈ2005ЃЉБЈЕРЕФКЌСПЃЌПЩФмЪЧвђЮЊБОЪЕбщЕФбјжГЫЎЮТЯрЖдНЯИпЧвВнгуЩњГЄНЯПьЃЌШЋГЬЮДГіЯжбсЪГЯжЯѓЁЃ
ЁЂ
ДгБэ2жаПЩМћЕААзжЪаЇТЪдк50 g/kgЫЧСЯжЌжЪзщжаДяЕНзюДѓжЕ2.69ЃЌдк75 g/kgКЭ100 g/kgЫЧСЯжЌжЪзщБЃГжЮШЖЈЃЌМфНгжЇГжСЫЪЪЕБЕФжЌЗОКЌСППЩНкдМЫЧСЯЕААзЕФРэТлЃЈDiasЕШЃЌ1998ЃЉЁЃДЫЭтЃЌЙ§СПЕФЫЧСЯжЌжЪЛсЕМжТВнгуЬхжЌЕФГСЛ§ЁЃВнгуШЋгужЌЗОКЌСПгыЫЧСЯжЌжЪКЌСПЕФе§ЯрЙиЙиЯЕЃЈБэ3ЃЉБэУїЃЌЕБЫЧСЯжЌжЪКЌСПГЌЙ§ВнгуашвЊСПКѓЃЌвЛВПЗжжЌжЪЛсзЊЛЏГЩШЋгужЌЗОГСЛ§ЯТРДЁЃЭЌбљЕФНсЙћдкЦфЫќгужжжавВгаБЈЕРЃЌШчКчїЎЃЈLeeЕШЃЌ1973ЃЉЁЂАпЕуВцЮВѕtЃЈGarlingЕШЃЌ1977ЃЉЁЂР№гуЃЈTakeuchiЕШЃЌ1979ЃЉЁЂКьЙФгуЃЈEllisЕШЃЌ1991ЃЉКЭдгНЛКњіѓгуЃЈJantrarotaiЕШЃЌ1994ЃЉЁЃ

4.2 ЫЧСЯжЌжЪЫЎЦНгыУтвпКЭНЁПЕ
дкЙ§ШЅШ§ЪЎФъЕФбаОПжаЗЂЯжЫЧСЯгЊбјГЩЗжАќРЈвЛаЉЬэМгМСПЩДЬМЄгуРрУтвпЯЕЭГВЂЬсИпПЙВЁФмСІЃЈViswanathЃЌ2012ЃЉЁЃдкБОЪЕбщжаЃЌЩшМЦСЫ5жжЕШЕЊЬнЖШжЌжЪЫЧСЯРДЦРЙРВнгуПЙИаШОПЙВЁФмСІЃЌНсЙћБэУїЫЧСЯжЌЗОЛсгАЯьВнгуУтвпгІД№ЁЃ
дкгуРрЕжПЙМВВЁжаЃЌЗЧЬивьадУтвпБШЬивьадУтвпИќживЊЃЈAndersonЃЌ1992ЃЉЁЃЭЬЪЩЯИАћвЛИіживЊЕФПЙОњгІД№ЪЧЛюадбѕЃЈROSЃЉВњЮяЃЈEoЕШЃЌ2008ЃЉЁЃБОЪЕбщНсЙћЯдЪОЭЬЪЩЯИАћЛюадбѕВњЮяЫцзХЫЧСЯжЌжЪКЌСПЕФдіМгЖјдіМгЃЌЕЋЪЧЕБЫЧСЯжЌжЪКЌСПГЌЙ§75 g/kgКѓЛсТдЮЂЯТНЕЃЈЭМ4AЃЉЁЃдкХЗжоіджавВЗЂЯжСЫЯрЭЌЕФЯжЯѓЃЈSitjЈЄ-BobadillaЕШЃЌ1999ЃЉЁЃШмОњУИБЛЙуЗКгУзїгуРрЕФУтвпжИБъЃЈOrtuneoЕШЃЌ2000ЃЉЁЃЫќгЩАзбЊЧђЗжУкВЂЗЂЛгживЊЕФПЙОњЛюадЁЃвдЭљЕФЪдбщжаШмОњУИЛюадКЭжЌжЪЕФВЙГфБэЯжГіСЫе§ЯрЙиЕФЙиЯЕЃЈLiЕШЃЌ2012ЃЉЃЌБОДЮЪЕбщжавВБэЯжГіСЫЯрЫЦЕФе§ЯрЙиЙиЯЕЃЈЭМ4CЃЉЁЃPODЪЧЯћГ§ЛюадбѕЕФПЙбѕЛЏУИЃЌвђДЫЕБгуРргІМЄКѓЦфPODЛюадЛсгаЫљдіМгЃЈRaicoЕШЃЌ2005ЃЉЁЃБОЪЕбщжаЩуЪГЮожЌЖдееЫЧСЯзщЕФВнгуPODЛюадзюИпЃЌЩуЪГ50~75 g/kgЫЧСЯжЌжЪзщЕФВнгуPODЛюадзюЕЭЃЌЕБВнгуИаШОЪШЫЎЦјЕЅАћОњКѓЃЌЫљгазщЕФPODЛюадМБОчдіМгЃЈЭМ4BЃЉЁЃИУНсЙћБэУїЮожЌЫЧСЯБШЙ§СПИпжЌЫЧСЯИќФмв§Ц№ВнгуЛњЬхгІМЄЗДгІЁЃ
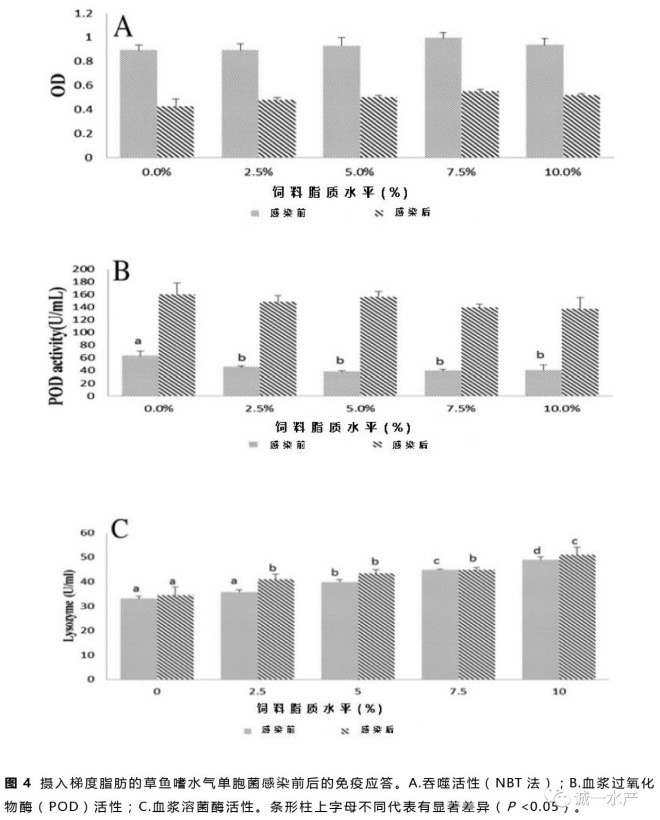
гЊбјбјжГЪЕбщКѓГЃГЃЭЈЙ§ВЁОњИаШОЪЕбщРДЦРЙРгуЬхЕФНЁПЕЃЈLimЕШЃЌ2009ЃЉЁЃЪЕМЪбјжГжаГЃМћЕФЪШЫЎЦјЕЅАћОњОГЃБЛгУдкгЊбјУтвпЪЕбщжаЃЌИаШОЪШЫЎЦјЕЅАћОњКѓЛсГіЯжГібЊадАмбЊжЂзДМАРЃбёадВЁБфЁЃдкЪЕМЪбјжГжаЪШЫЎЦјЕЅАћОњИаШОвВЪЧЕЫЎгубјжГИпЫРЭіТЪЕФдвђжЎвЛЃЌдьГЩСЫОоДѓЕФОМУЫ№ЪЇЃЈJiravanichpaisalЕШЃЌ2009ЃЉЁЃШчЭМ3ЫљЪОЃЌдкЪШЫЎЦјЕЅАћОњИаШОКѓЃЌЮожЌЖдеезщВнгуЕФДцЛюТЪзюЕЭЃЌЫЕУївћЪГжаЕФжЌжЪвЛЖЈГЬЖШЩЯЬсИпСЫПЙВЁФмСІЁЃдкдчЦкХЗжоКЃідМАЮфВ§гуЕФЪЕбщжаЖМГіЯжСЫвЛжТЕФНсЙћЃЈLiЕШЃЌ2012ЃЉЁЃвВгабаОПБЈЕРБэУїЫЧСЯжЌжЪЫЎЦНЖдТоЗЧгуПЙСДЧђОњЃЈS. iniaeЃЉИаШОУЛгагАЯьЃЈLimЕШЃЌ2009ЃЉЃЌдкАпЕуВцЮВѕtЩуЪГВЛЭЌжЌжЪЫЎЦНЃЈ0~90 g/kgЃЉЫЧСЯКѓНјааЕФАЎЕТЛЊЪЯОњИаШОЪЕбщжаЃЌНсЙћвВУЛгаГіЯжВювьЃЈYildirim-AksoyЕШЃЌ2009ЃЉЁЃЪЕбщНсЙћЕФВювьПЩФмЪЧгЩгкЪЕбщгужжВЛЭЌЁЂДѓаЁВЛЭЌЁЂЫЧСЯжЌжЪдДВЛЭЌЁЂЭЖЮЙТЪВЛЭЌЛђбјжГЬѕМўВЛЭЌЕШдвђдьГЩЃЈLiЕШЃЌ2012ЃЉЁЃдкДѓЮїбѓійгуЕФЯрЙибаОПжаЗЂЯжЩуЪГгугЭЕФгуБШЩуЪГВЫзбгЭЕФгудкГйЛКАЎЕТЛЊЪЯОњИаШОЪЕбщжаЕФДцЛюТЪИќИпЃЈCarterЕШЃЌ2003ЃЉЁЃдкДѓЮїбѓійгугУПћЛЈгЭВЛЭЌГЬЖШЕФЬцДњгугЭЪЕбщжаЃЌГйЛКАЎЕТЛЊЪЯОњИаШОКѓРлЛ§ЫРЭіТЪГіЯжСЫЯджјВювьЃЈBransdenЕШЃЌ2003ЃЉЁЃАпЕуВцЮВѕtЕФвЛИідчЦкЪЕбщжИГіЫЧСЯжЌжЪЃЈіюгугЭКЭбЧТщзбгЭЃЉдкИпЮТЧщПіЯТЃЈ28ЁцЖдБШ17ЁцЃЉгАЯьАЎЕТЛЊЪЯОњИаШОКѓЕФЫРЭіТЪЃЈFracalossiЕШЃЌ1994ЃЉЁЃ
БОЪЕбщжаЃЌдкЪШЫЎЦјЕЅАћОњИаШОЯТЩуЪГ75 g/kgЫЧСЯжЌжЪзщЕФВнгуДцЛюТЪзюИпЃЌЕБЫЧСЯжЌжЪЫЎЦНЩЯЩ§ЕН100 g/kgЪБЃЌДцЛюТЪгаЫљЯТНЕЃЈЭМ3ЃЉЃЌБэУїЙ§СПЕФЫЧСЯжЌжЪВЛРћгкВнгуЗЧЬивьадУтвпЯЕЭГЃЌПЩФмЪЧгЩгкЙ§СПЕФжЌжЪЖдИЮдрдьГЩСЫЦЦЛЕЁЃ
ЙШВнзЊАБУИКЭЙШБћзЊАБУИЪЧгуЬхФкживЊЕФзЊАБУИЃЌГЃБЛгУзїгуРрНЁПЕжИБъЃЌгааЇЬхЯжИЮдрЕФЪмЫ№зДЬЌЃЈSheikhzadehЕШЃЌ2012ЃЉЁЃБОЪЕбщжаЃЌбЊНЌЙШВнзЊАБУИКЭЙШБћзЊАБУИЛюаддкЩуЪГ50~75 g/kgЫЧСЯжЌжЪзщЕФВнгужазюЕЭЃЈБэ4ЃЉЃЌБэУїЫЧСЯжЌжЪЫЎЦНдк60 g/kgзѓгвЪБЖдВнгуИЮдрЙІФмЕФгАЯьзюаЁЁЃДЫЭтЙШВнзЊАБУИКЭЙШБћзЊАБУИЛюаддкЩуЪГЮожЌЖдееЫЧСЯКЭ100 g/kgжЌжЪЫЎЦНЫЧСЯЕФВнгужаЯджјЩ§ИпЃЌЫЕУїЮожЌЛђИпжЌЖМЛсвђЮЊМгжиДњаЛИКЕЃЖјгАЯьВнгуИЮдрЕФе§ГЃЙІФмЃЈHevrøyЕШЃЌ2004ЃЉЁЃЭЌбљЕФНсЙћдкжЎЧАЕФВнгугзгуЪЕбщЃЈLinЕШЃЌ1990ЃЉМАЮфВ§гуЪЕбщЃЈLiЕШЃЌ2012ЃЉжавВгаЗЂЯжЁЃ

бЊНЌМюадСзЫсУИЛюадЭЌбљБЛгУгкЦРЙРИЮдрЕФЫ№ЩЫГЬЖШЃЈVenkateswaraЃЌ2006ЃЉЁЃЕБИЮдрМюадСзЫсУИЛюадНЕЕЭЛђбЊНЌМюадСзЫсУИЛюадЩ§ИпЖМПЩзїЮЊИЮдрБЁБкзщжЏЕФЫ№ЩЫЕФБъжОжЎвЛЁЃБОЪЕбщжаЃЌбЊНЌМюадСзЫсУИЛюаддкЮожЌЖдеезщжазюИпЃЈБэ4ЃЉЃЌБэУїЮожЌЫЧСЯЖдИЮдрЕФЫ№КІПЩФмИпгкИпжЌЫЧСЯЁЃМюадСзЫсУИЛюадЫцзХЫЧСЯжЌжЪЫЎЦНЩ§ИпЖјЯТНЕПЩФмЪЧгЩгкВнгуж№НЅЪЪгІСЫИпжЌЫЧСЯЃЈBanaeeЕШЃЌ2011ЃЉЛђепзЊСзЫсЛюадЯТНЕЃЈSharmaЃЌ1990ЃЉЁЃ
бѕЛЏгІМЄЭљЭљЪЧвђЮЊЛюадбѕВњЮяЕФВњЩњКЭЯћГ§ВЛЦНКтдьГЩЕФЃЈTocherЕШЃЌ2002ЃЉЁЃвЛАуРДЫЕЃЌгуРрЕФПЙбѕЛЏЗРЛЄЪмгЊбјвђзгЕФгАЯьНЯДѓЃЈSheikhzadehЕШЃЌ2012ЃЉЁЃБОЪЕбщжаЃЌИЮдрБћЖўШЉЃЈMDAЃЉКЌСПЫцзХЫЧСЯжЌжЪКЌСПЕФЩ§ИпЖјЛКТ§ЩЯЩ§ЃЈЭМ2AЃЉЃЌБэУїВнгуЖджЌЗОЫсЙ§бѕЛЏЗДгІЕФУєИаадЩЯЩ§ЃЈStЈІphanЕШЃЌ1995ЃЉЁЃдкВнгуЩуЪГВЛЭЌжЌЗОдДЕФИпжЌЫЧСЯжаЃЈDuЕШЃЌ2008ЃЉМАЮфВ§гуЩуЪГВЛЭЌЬЧжЌБШЕФЫЧСЯЪЕбщжаЃЈLiЕШЃЌ2012ЃЉвВГіЯжСЫЯрЫЦНсЙћЁЃЯрЗДЃЌзмПЙбѕЛЏУИЃЈT-AOCЃЉЛюадЫцзХЫЧСЯжЌжЪЫЎЦНЕФЬсИпЖјЯджјНЕЕЭЃЈЭМ2CЃЉЃЌБэЯжГіМССПвРРЕадЃЌЫЕУїВнгуПЙбѕЛЏФмСІЯджјЪмЫЧСЯжЌжЪКЌСПЕФгАЯьЧвT-AOCЛюаддкНёКѓЕФбаОПжаПЩзїЮЊгуРрУтвпгІД№ЕФжИБъжЎвЛЁЃЫфШЛИїзщМфИЮдрГЌбѕЦчЛЏУИЃЈSODЃЉЛюадУЛгаЯджјадВювьЃЌЕЋЪЧЩуЪГНЯИпЫЧСЯжЌжЪЕФВнгуИЮдрSODЛюадГіЯжЩЯЩ§ЕФЧїЪЦЃЈЭМ2BЃЉЁЃSODзіЮЊвЛжжПЙбѕЛЏУИПЩДпЛЏСНИіГЌбѕздгЩЛљОЙ§ЦчЛЏзїгУБфГЩЙ§бѕЛЏЧтКЭбѕЦјРДЯћГ§Й§бѕЛЏЮяЃЈFattmanЕШЃЌ2003ЃЉЁЃзмЕФРДЫЕЃЌБОЪЕбщжаИЮдрбѕЛЏМАНЁПЕжИБъMDAЁЂSODКЭT-AOCЫљБэЯжГіРДЕФЧїЪЦБэУїИпжЌвћЪГЕФВнгубѕЛЏГЬЖШИпЁЃ
БОЪЕбщжабЊНЌИЪгЭШ§ѕЅКЭзмЕЈЙЬДМКЌСПЫцзХЫЧСЯжЌжЪЫЎЦНЕФЩ§ИпЖјЩ§ИпЃЈБэ4ЃЉБэУїЫЧСЯжЌжЪЫЎЦНЕФЬсИпЪЙЕУВнгуФкдДаджЌЗОзЊдЫИќЛюдОЁЃИУНсЙћдкжЎЧАЕФбаОПжавВгаРрЫЦБЈЕРЃЈDuЕШЃЌ2005ЃЉЃЌБэУїИпЫЧСЯжЌжЪЫЎЦНПЩФмМгжиИЮдрДњаЛИКЕЃЁЃ
5ЁЂНсТл
ДгБОЪЕбщНсЙћПЩжЊЪЪСПЕФЫЧСЯжЌжЪЫЎЦНПЩДйНјВнгуЛњЬхУтвпФмСІЃЌЕЋЪЧЙ§СПЕФЫЧСЯжЌжЪЫЎЦНЛсМгжиИЮдрбѕЛЏГЬЖШЁЃДгЪЕгУНЧЖШПДЃЌБОЪЕбщНсЙћНЈвщдке§ГЃЕФбјжГзДЬЌЯТВнгуЫЧСЯжЌжЪКЌСПвд65.3 g/kgЮЊМбЃЌЕЋЪЧдкЬиЪтЪБЦкгШЦфЪЧМВВЁвзЗЂЪБЦкЃЌЪЪЕБЬсИпЫЧСЯжЌжЪКЌСПЪЧЪЎЗжБивЊЕФЁЃИУЪБЦкПЩНЋжЌЗОКЌСПЬсИпЕН75 g/kgРДЬсИпВнгуПЙгІМЄФмСІЁЃЃЈЭъЃЉ
1ЁЂРДдДЃКЁЖжаДѓЫЎЩњЭЈбЖЁЗЕк59Цк
2ЁЂзїепЃКжаЩНДѓбЇЩњУќПЦбЇбЇдКЫЎЩњОМУЖЏЮябаОПЫљ Н№бо/ЮФ
3ЁЂЮЂаХКХЃКgzchengyi2013



